
Polyploids are defined as organisms with other than 2 basic sets of chromosomes, that is, monoploids, triploids, tetraploids, and various aneuploids. Segregation in polyploids differs from that in diploids due to the many different ways in which the chromosomes can pair in a polyploid and due to larger number of alleles segregating at each locus.
Judging by the frequency with which different kinds of polyploids appear among crop plants, autotetraploidy appears to be more important to agriculture than the others. Four of the major crop species, namely, alfalfa, potato, coffee and peanut may be autotetraploids. In addition, several forage grasses and many ornamentals appear to be autotetraploids.
In respect of one locus with 2 alleles, A and a, 5 genotypes are possible in an autotetraploid. These are AAAA, AAAa, AAaa, Aaaa and aaaa. According to the number of dominant alleles, these 5 genotypes are referred to as quadruplex, triplex, duplex, simplex, and nulliplex respectively.
The meiosis in autotetraploids involves the partitioning of the 8 chromatids during meiosis I into 4 pairs, each pair corresponding to one of the 4 gametes produced by each sporocyte.
This process is much more complicated than that of diploids where meiosis involves the partitioning of only 4 chromatids, one to each of the 4 gametes. Depending upon the regularity of quadrivalent formation and the variable chiasma number between the kinetochore and the locus in question, 2 kinds of segregations are possible in the autotetraploids.
Random Chromosome Segregation:
The type of partitioning of chromatids where sister chromatids i.e., the chromatids derived from the same chromosome never end in the same gamete is called as random chromosome segregation. This happens only when bivalents are formed. For example, in a simplex (Aaaa) the segregation of chromosomes during meiosis is as follows. Here 8 chromatids are shown as (A1 A2), (a3 a4), (a5 a6) and (a7 a8).
The gametic output from any autotetraploid can be easily derived by making all possible pairs of chromosomes out of 4 chromosomes as given below:

A checkerboard method of deriving the gametes under random chromosome assortment for a duplex is shown in Fig. 5.5:

The Fig. 5.5 shows that two sister chromatids i.e., the chromatids derived from the same chromosome, for example, A1 and A2 and similarly other ones are not included in the same gamete as required under random chromosome assortment. The gametic output from the checkerboard is 4 AA : 16 Aa : 4 aa i.e. 1 A A : 4 Aa : 1 aa.
The zygotic frequencies can be easily worked out as given below:
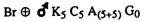
Likewise various other gametic and phenotypic ratios can be determined.
Random Chromatid Segregation:
When quadrivalent formation is complete and there is occurrence of 50% crossing over between centromere and locus, the partitioning of 8 chromatids to the gametes is at random i.e., all the possible pairs of 8 chromatids of an autotetraploid have an equal chance of being included in the same gamete following meiosis. The gametic ratio under this system for a triplex (AAAa) as an example is given in Fig. 5.6. The gametic ratio is 15 AA : 12 Aa : 1 aa.

Given random pairing and 50% crossing over and tracing the relationship of 8 chromatids to the 4 centromeres in every spore mother cell and then through meiosis I and meiosis II, the gametic outputs are slightly different than under random chromatid segregation.
For example, a triplex yields gametes in a ratio of 15 AA : 12 Aa : 1 aa under chromatid segregation, but a ratio of 13 AA : 10 Aa : 1 aa under 50% crossing over situation.

It must be kept in mind that random chromosome assortment and random chromatic assortment are two theoretical extremes and real gametic output is intermediate between these extremes depending upon whether quadrivalents form or do not form and when centromere and the locus are partly linked.
The net effect of these 2 events is reflected in the value of alpha which itself is related to gametic output, for example, for a simplex, the gametic expectations in terms of alpha are:
![]()
For random chromosome assortment, α = 0 and for random chromatid assortment α = 0.1429. In an actual situation it is in between these two extreme values. Table 5.5 shows that the frequency of recessive phenotypes is much lower than in the equivalent diploids. Thus, auto-polyploids are genetically more stable than diploids and release their allelic variability at a much slower rate.