
The below mentioned article provides a short note on the Homeotic Genes.
Homeotic genes constitute an intricate regulatory network, in which one gene may activate or repress other genes Homeotic genes are involved in regulation of development through a predetermined pathway in which specific genes are turned on and off at particular times.
Homeotic genes act in a transcriptional cascade, in which a series of hierarchical interactions between the regulatory proteins is succeeded by the activation of structural genes coding for morphogenesis.
Homeotic genes are situated on a particular segment of chromosome, may be as multi-gene or as cluster of genes which interact in a complicated interlocking pattern that is not fully defined. Many homeotic genes code for transcription factors that act upon other homeotic genes as well as upon other target loci. As a result, a mutation in one homeotic gene may influence the expression of other homeotic genes.
The molecular organization of some homeotic genes is unusual and gave rise to their description as complex loci, and itself poses some questions about gene expression. The homeotic mutants develop organs at wrong places due to inability to recognise their positional information properly, as a result the organs themselves were normal but their positions at which they developed were improper.
Embryogenesis in Drosophila:
Homeotic genes are involved during embryogenesis in Drosophila. Their expression depends on the prior expression of segmentation genes.
The segmentation genes define the number and locations of segments and homeotic genes impose the programme that determines the unique differentiation of each segment. So homeotic mutants cause the transformation of a segment to develop phenotype of other segment, i.e., legs may develop in place of antennae or wings may develop in place of eyes, etc.
The classic complex homeotic locus is BX-C, the bithorax complex, characterized by several groups of homeotic mutations that affect development of the thorax, causing major morphological changes in the abdomen (Fig. 14.26). Another such example is ANT-C (antennapedia complex), affecting head and thorax.
These homeotic genes, which were also called Hox genes, are characterized by the presence of a homeobox of 180 nucleotide sequence, translated into 60 amino acids domain called homeodomain. These are actually DNA binding protein with helix-turn-helix (HTH) conformation playing regulatory role for transcription of DNA by binding at major groove of DNA double helix.
However, Hox genes represent only a subset of homeobox genes, since-many genes containing a homeobox may not be homeotic in nature.

Flower Primordia Development:
Induction of flower development in plants takes place by the changes from vegetative to reproductive meristems or floral primordia and homeotic genes act sequentially controlling development. Flower development requires a cascade of sequential gene activity that gradually converts a mass of undifferentiated apical meristem cells into the different parts of a flower.
The genes encode transcription factors that act as master switches turning on or off downstream genes that ultimately make each part of the flower in its appropriate location.
The mechanism of flower morphogenesis in Arabidopsis has been deduced through a genetic model called ABC model.
The floral homeotic genes have been studied and classified into 3 categories – A, B and C. Genes from category A – APETALA1 (AP1) and APETALA2 (AP2) are expressed in sepal and petal primordia; genes from category B – APETALA3 (AP3) and PISTILLATA’ (PI) are expressed in petal and stamen primordia; and gene from category C – AGAMOUS (AG) affect stamens and carpels (Fig. 14.27).
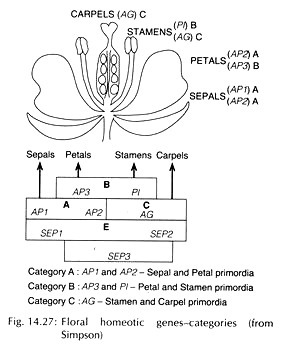
According to the revised ABCE model, another category (E) of genes – SEPALLATA (SEP1-4) act in combination with A, B, C genes for specification of floral organs. The LEAFY (LEY) gene identified, is responsible for specifying floral meristem identity and plays a major role in turning on the above groups of genes for floral organ development.
After expression of the gene AGAMOUS (AG), involved in the development of stamen and carpel, it is repressed by the repressor which is another gene product of CURLY LEAF.
In floral development, the class A gene and class C gene are believed to function antagonistically, because, in a mutant (ap2) of class A gene, the expression of class C (AG) gene expanded to whorls one and two. In mutant (ag) of class C gene, AP1 expands its expression to the inner whorls.
The expression of PI and AP3, i.e., class B genes is independent of class A and C, but PI activity is required for AP3 expression and vice versa (Table 14.2).

Homeotic genes have also been identified in other plant systems like Antirrhinum (LIP, DEF, GLO, PLE, FAR), Petunia (GP, BL, DDL, PH3), Gerbera (GRCD), etc. These floral organ identity genes have been shown to encode transcription factor each with conserved MADS-DNA binding domain.
MADS domain has 56 amino acids which was initially found in 4 different proteins: transcription factor MCM of yeast, proteins encoded by AGAMOUS and DEFICIENS genes of Arabidopsis and Antirrhinum, transcription factor of SRF of mammals; and from these four proteins the name ‘MADS’ has been originated.
This MADS domain is a DNA binding domain and also helps in protein-protein interactions. The homeotic MADS box genes are actually a subset of a larger MADS box gene family, some of whose members are expressed in all tissues, whereas others show tissue or organ specific expression.
Many MADS box genes have been cloned from various plants and expression patterns show the common gene regulatory system in flower development. De Folter et al. (2005) have concluded that self-regulation is a re-occurring phenomenon among the MADS domain proteins.
Since many single MADS domain proteins can form complexes with several other MADS domain proteins, the number of possible combinations is very large and may permit the proteins to function in different contexts. The functional view of, for example, the AG protein, in determining stamen and carpel identity, is that AG interacts with different sets of MADS domain factors to carry out the different functions.